
1 引言
人们在理解他人动作时,不仅预判动作的空间结果,也会预判动作的时间结果(Diersch et al., 2016)。例如,在接住别人抛来的盒子时,人们除了判断盒子落下的位置,也会预估落下的速度。因此,速度预判是在物体移动前或移动初期对其经过特定点时速度的提前判断。同时,在动作理解的范畴下,速度预判是动作预判(action anticipation)在时间维度上的加工,是时间知觉的表现方式(Manning et al., 2018)。在充满人际互动的体育场上,尤其需要精确的速度预判来完成各项拦截动作。目前,已有大量的研究证明了运动专家对熟悉动作的身体运动学感知或模拟能力更强,能够对动作预判的空间结果判断更加准确(Diersch et al., 2016; Pedullà et al., 2020; Vicario et al., 2017)。然而,运动经验是否也能够促进其对动作时间的模拟,表现出更强的速度预判,仍缺乏系统的研究。
根据模拟理论,人们对他人动作的理解是通过所观察动作的视觉表征,映射到同一动作的运动表征上而实现的(Hesslow, 2012)。也就是说,相比没有运动经验的群体,运动专家可以通过视觉提取运动学信息,利用运动表征,准确模拟出动作结果。大量研究也验证了运动专家能够通过模拟动作准确预判落点位置(Baurès et al., 2018),但在时间维度的模拟准确性上并没有得到一致的结果。以往研究利用时间遮蔽范式,通过判断遮蔽后出现的动作是否具有时间连贯性,来考察个体对动作的速度预判加工(Stadler et al., 2012)。研究发现,当呈现的遮蔽后动作时间连贯性越高,判断的正确率就越高(Vicario et al., 2017)。然而,也有研究发现,对动作速度的模拟可能会快于实际动作(Stadler et al., 2012),也可能会慢于实际动作(Brich et al., 2018)。这些研究提示了动作理解中的速度预判可能同时受对运动学信息的模拟能力和对物体移动速度的感知能力影响,也就是说物体本身的移动速度会干扰遮蔽过程中的动作速度模拟。
在针对运动情境中的速度预判研究发现,花样滑冰运动员会比普通人更精准地进行速度预判,提示了运动经验对速度预判的促进作用(Diersch et al., 2012)。而在预判篮球投篮成功性的任务中,篮球运动员在延迟出现动作结果的遮蔽条件下有更好的判断,推测运动专家在动作表征过程中通过放慢表征速度实现更精细的模拟过程(Vicario et al., 2017)。这些结果提示了运动经验对速度预判的促进可能依赖于对动作意图的理解。根据动作理解的层级理论,对动作理解过程既可以发生在运动学信息的低层级模拟,也可以产生动作意图的高层级理解(赵峰 等, 2016)。前人在与速度预判密切相关的足球拦截时机判断任务中发现,对拦截时机的预判加工不仅依赖于相关的运动经验,也包含对动作意图的理解(Pedullà et al., 2020)。因此,本研究推测基于动作理解的速度预判可能依赖于高层次的动作意图加工,在表征过程中可能会激活动作理解的脑区。
动作观察网络(action observation network, AON)是个体在观看和理解熟悉动作时激活的主要脑区,包括前运动皮层、颞上沟、顶下小叶(inferior parietal lobule, IPL)、额下回等关键节点(Abreu et al., 2012)。其中IPL被认为是参与理解动作意图的重要节点(Rizzolatti & Sinigaglia, 2010)。相比一般认知任务,羽毛球运动员通过强化运动皮层与右侧IPL的连接来完成动作预期(Xu et al., 2016)。对动作意图的理解会激活除运动系统之外与语义加工有关的枕颞区(Wurm & Lingnau, 2015)。枕颞皮层在动作预期的时间维度加工中也发挥作用(Diersch et al., 2016)。这些研究提示了顶叶区域在识别经验信息中的关键性,而且顶叶和颞叶区域能够编码感知上的不同特征,从而能够区分具有不同动作目标的动作(Lestou et al., 2008)。基于这些发现,本研究预测,速度预判需要依赖动作意图的理解。因此,相比无经验人群,运动专家在观看动作进行速度预判时会激活AON相关脑区,并且IPL和枕颞区可能是表征动作速度的重要脑区。
本研究通过对比足球运动员和普通大学生对点球动作的速度预判加工过程的行为表现和大脑活动,探讨运动专家在专项动作速度预判加工中的认知优势。本研究通过2个行为实验和1个功能磁共振成像(fMRI)实验展开。首先,通过动作预判的速度一致性辨别任务验证足球运动员具有速度预判的专家优势。其次,利用时间阻断技术,进一步验证足球运动员速度预判加工中对运动学信息的利用特征。最后,通过fMRI技术结合单变量和多体素模式分析(MVPA),验证动作意图理解的关键节点在速度预判加工中的作用。本研究预期具有丰富运动经验的足球运动员在点球动作的速度预判中具有更好的速度变化辨别能力,并且加工过程更依赖运动学信息和动作意图的理解,IPL和枕颞区可能是参与速度预判的重要脑区。
2 实验1:足球运动员对点球动作速度预判加工优势
检验足球运动员(专家组)相比普通大学生(对照组)对足球点球动作具有速度预判的加工优势。根据模拟理论,足球运动员可以根据已有的运动经验模拟点球动作结果,当出现球速与踢球动作不连贯时,预期足球运动员对速度变化的辨别会更加敏锐。
2.1 被试
使用G*Power3.1估计研究中被试所需的样本量,本研究将效应量(effect size)设置为0.25,Power值(1−β)设置为0.95,α设置为0.05,重复测量次数4,计算出最小样本量为36人,即每组18人。为控制性别变量,本研究共招募30名女性专业足球运动员为专家组(国家一级运动员26人,健将4人),30名无足球运动经验的女大学生为对照组。根据利手问卷(Edinburgh Handedness Inventory),所有被试均符合右利手标准,且主观报告视力或矫正视力正常,无神经或精神疾病史。两组被试的人口统计学特征见表1 。所有被试在实验前均签署了知情同意书,实验结束后获得金钱报酬。本实验得到上海体育大学伦理委员会批准(102772023RT115)。
表1 实验1被试基本信息(M±SD) |
| 变量 | 专家组 | 对照组 | t | p |
| 样本量 | 30 | 30 | ||
| 身高(cm) | 166.60±5.21 | 164.53±5.84 | 1.45 | 0.162 |
| 体重(kg) | 57.10±6.18 | 54.50±9.51 | 1.26 | 0.213 |
| BMI(kg/m²) | 20.61±1.52 | 20.29±2.85 | 0.54 | 0.591 |
| 年龄(岁) | 20.97±1.69 | 20.43±2.22 | 1.05 | 0.302 |
| 训练年限(年) | 9.63±2.91 | 0 | 18.15 | <0.001 |
2.2 实验材料
刺激材料为自行录制的足球点球视频。录制模特为2名国家一级女子足球运动员(年龄22岁、23岁)。将球放置在国际足联规定的点球位置,从球的左后侧2米处进行跑动,初始位置与球的夹角为45°,执行右脚脚内侧推射动作。录制完整点球视频20组,两名运动员各10组,每组包含向左、向右射门方向的快球和慢球各一个。录像设备为富士T100(分辨率1280×720像素,60帧/秒),设备位于球门中点,正对点球放置点。确认入选快速球20段(平均球速23.45 m/s, 最慢速度为22.00 m/s),慢速球20段(平均球速17.80 m/s, 最快速度为19.00 m/s),模特各半、方向各半。
利用Adobe Premier(Adobe Systems Incorporated, 美国)对基础材料进行剪辑,将球与脚接触为关键帧,进行动作与球飞行片断的重新组合,形成一致(快踢球−快速球、慢踢球−慢速球)和不一致(快踢球−慢速球、慢踢球−快速球)两类刺激材料(图1 )。
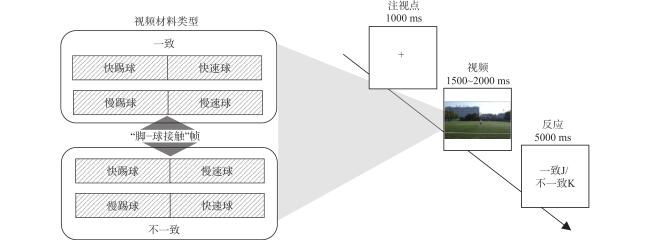
2.3 实验流程
采用2×2混合设计,组间变量为运动经验(专家组、对照组),组内变量为一致性(一致、不一致)。所有试次开始于屏幕中央1000 ms的注视点“+”,随后呈现1500~2000 ms点球视频,被试在视频播放后判断点球动作和球速的一致性,若判断一致,按“J”,若不一致,按“K”(图1 )。任务共包含16个练习试次和160个正式试次,正式试次中一致与不一致为1∶1。采用E-Prime3.0(Psychology Software Tools, 美国)呈现刺激。
2.4 数据采集与分析
采用E-Prime3.0进行行为数据的收集,记录被试的正确率和反应时。行为数据服从正态分布,使用SPSS24.0进行统计分析。分别对正确率和正确判断试次的反应时进行2(运动经验:专家组、对照组)×2(一致性:一致、不一致)混合设计方差分析。进行Mauchly’s Test检验球形假设,若球形假设不成立,使用Greenhouse-Geisser校正,事后多重比较采用Bonferroni校正。
2.5 结果
正确率结果显示,运动经验主效应显著,F(1, 58)=12.60,p<0.001,η${_{\rm p}^2} $ =0.18,专家组的正确率(0.65±0.01)显著高于对照组(0.60±0.01)。一致性主效应显著,F(1, 58)=427.51,p<0.001,η${_{\rm p}^2} $ =0.89,一致的正确率(0.74±0.02)显著高于不一致(0.52±0.02)。运动经验和一致性交互作用不显著,F(1, 58)=2.70,p=0.900。反应时结果显示未发现显著差异。
2.6 讨论
实验1结果表明,专家组能够更好地识别出踢球动作与动作结果之间的一致性,具备更好速度预判加工能力。这一结果符合模拟论,运动专家能够准确地根据所提取的运动学信息,借助内部模型对当前动作进行表征,从而更好进行速度预期加工。并且,无论呈现的动作结果是否与运动学信息匹配,专家组的整体预判正确率都优于对照组,由此推测足球运动员的速度预期优势并不会受到时间连贯性的影响。对于不一致,两组群体均表现出的正确率下降,可能是由于相对识别一致结果,不一致结果的识别难度更高,这也与其他运动项目在空间预判上的结果一致(Vicario et al., 2017)。
3 实验2:足球运动员对点球动作速度预判的运动学信息加工
实验2采用时间阻断技术探索专家速度预判优势的信息加工特征,推测摆腿动作是足球运动员进行速度预判的关键运动学信息。
3.1 被试
与实验1采用相同被试库,其中18名运动员(19.06±0.80岁)和19名普通大学生(19.26±0.81岁)完成实验2。
3.2 实验材料
对基础材料进行时间阻断剪辑,分别形成助跑(900 ms)、摆腿(800 ms)和从助跑到踢出球的控制条件(1500 ms)三种运动学信息类型的刺激材料。
3.3 实验流程
采用2×3混合设计,组间变量为运动经验(专家组、对照组),组内变量为运动学信息类型(助跑、摆腿、控制条件)。所有试次开始于屏幕中央1000 ms的注视点“+”,随后呈现800~1500 ms的视频材料,被试在视频结束后预判快球按“F”,慢球按“G”(图2 )。实验共包括12个练习试次和240个正式试次,每种运动学信息类型各80试次,时长约20分钟。
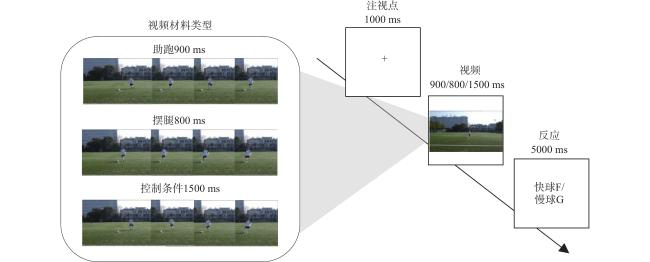
3.4 数据采集与分析
同实验1。
3.5 结果
正确率结果显示,组别主效应显著,F(1, 33)=11.80,p=0.002,η${_{\rm p}^2} $ =0.25,专家组的正确率(0.75±0.01)显著高于对照组(0.69±0.01);运动学信息类型主效应显著,F(2, 66)=47.27,p<0.001,η${_{\rm p}^2} $ =0.74,在控制条件正确率(0.79±0.01)显著高于助跑(0.70±0.01)和摆腿(0.68±0.01),助跑与摆腿无显著差异;交互作用不显著,F(2, 66)=2.77,p=0.080。
反应时结果显示,运动学信息类型主效应显著,F(2, 66)=15.19,p<0.001,η${_{\rm p}^2} $ =0.47,控制条件反应时(519±27 ms)显著快于助跑(555±24 ms)和摆腿(646±33 ms),助跑显著快于摆腿,组别主效应[F(1, 33)=1.84, p=0.180]和交互作用不显著[F(2, 66)=1.27, p=0.300]。
3.6 讨论
结果提示,专家组速度预判的整体正确率表现优于对照组,这与前人空间预判结果一致(Xu et al., 2016)。但是,观看助跑和观看摆腿的速度预判表现没有差异。反应时结果发现,摆腿比助跑有更长的速度预期加工时间。依据动作加工的层级理论推测,摆腿的加工相比助跑需要更多的加工过程,这可能是由于助跑的加工属于低层级的运动学信息加工,倾向于通过速度感知预判动作结果;而通过摆腿的速度预判属于高层级的抽象加工,倾向于通过意图理解来预判动作的速度结果。实验3借助fMRI技术,通过对点球动作速度预判过程中的大脑激活来验证这一推测。
4 实验3:足球运动员速度预判的大脑加工特征
实验3比较运动员和普通大学生在速度预判中的大脑活动,利用MVPA解码速度验证加工速度预期的关键脑区。
4.1 被试
同实验1。
4.2 实验材料
选择剪辑完成的刺激材料20段,其中快速和慢速各半。每段视频均包含完整的助跑到球踢出过程,视频时长2000 ms。
4.3 fMRI流程
被试需要观看足球点球视频,判断踢出的球是快速还是慢速,在正式实验前,先在核磁外电脑上进行练习(20个试次)。正式实验采用event设计,共160个试次,快速和慢速各半,时长约12分钟。所有试次开始于屏幕中央1000 ms的注视点“+”,随后呈现2000 ms的点球视频,被试在视频播放的同时即可做出按键反应,判断快速食指按键,慢速中指按键。
4.4 数据分析
4.4.1 fMRI数据采集和预处理
采用西门子Prisma 3.0T磁共振扫描仪(3.0 T Siemens Trio)和58通道头线圈。BOLD功能像序列,采用梯度回波平面成像序列(echo planar imaging, EPI)采集,重复时间(repetition time, TR)=2000 ms,回波时间(echo time, TE)=30 ms,翻转角(flip angle, FA)=90°,层数(slice)=58,视野大小(field of vision, FOV)=192 mm×192 mm,体素大小(voxel size)=2 mm×2 mm×2 mm。T1加权结构像参数:TR=2530 ms,TE=2.98 ms,FA=7°,slice=192,voxel size=1 mm×1 mm×1 mm。被试仰卧在扫描仪内,要求在扫描过程中保持头部及身体静止。
实验所获取的图像数据采用加载于MATLAB2017b平台上的SPM12软件包进行预处理及统计分析。预处理中为了提高磁场均匀性及排除被试对核磁环境的适应过程,去掉扫描模块的前10张图像,之后进行时间层校正以及头动校正,排除头部平动大于2 mm、转动大于2°的被试。在空间标准化中,首先将每个被试的结构像配准到平均功能像中,之后将功能像标准化到标准脑模板空间(Montreal Neurological Institute, MNI),最后采用高斯平滑核(FWHM=8 mm)进行空间平滑。
4.4.2 fMRI数据统计分析
首先对所有数据进行全脑分析,利用SPM12对速度预期过程建立一般线性模型(GLM),同时为了避免轻微头动带来的信号失真,将被试的头动参数作为协变量进行回归。脑数据体素水平未校正,p<0.001,团块水平的校正阈值设置为FDR校正,p<0.05。
其次基于ROI对脑成像数据进行分析,选取双侧的IPL(左: −32, −48, 60; 右: 32, −48, 60)、额下回(inferior frontal gyrus, IFG; 左: −48, 6, 18; 右: 48, 12, 16)、背外侧前额叶(dorsolateral prefrontal cortex, DLPFC; 左: −52, 8, 24; 右: 58, 8, 30)以及外侧枕颞皮层(lateral occipitotemporal cortex, LOTC; 左: −36, −54, −14; 右: 36, −54, −14)作为ROI,通过相关研究中激活的峰值坐标,以该坐标为中心,建立以10 mm为半径的小球作为ROI(Schubotz & von Cramon, 2009)。采用MarsBaR0.45提取ROI的BOLD信号变化百分比值,使用SPSS24.0对信号值进行独立样本t检验和配对样本t检验。
最后使用Python3.10的Nilearn(Abraham et al., 2014)对专家组和对照组在完成任务时的大脑激活模式进行了MVPA。对于每个ROI内的体素,在数据子集上训练了一个支持向量机(SVM)分类算法,采用5折交叉验证法训练SVM分类器进行对速度快慢编码的大脑活动模式分类(Stelzer et al., 2013)。
4.5 结果
4.5.1 行为任务结果
正确率的组别主效应显著,F(1, 58)=17.50,p<0.001,η${_{\rm p}^2} $ =0.23,专家组的正确率(0.80±0.02)显著高于对照组(0.67±0.02);速度类型主效应显著,F(1, 58)=5.76,p=0.020,η$ {_{\rm p}^2}$ =0.09,对快速球预判的正确率(0.77±0.02)显著高于慢速球(0.70±0.02);交互作用不显著。反应时的组别主效应显著,F(1, 58)=4.62,p=0.040,η${_{\rm p}^2} $ =0.07,专家组的反应时(1683±37 ms)显著快于对照组(1794±37 ms);速度类型主效应显著,F(1, 58)=11.22,p=0.001,η${_{\rm p}^2} $ =0.16,对慢速球预判的反应时(1716±25 ms)显著快于快速球(1761±28 ms),交互作用不显著,F(1, 58)=2.296,p=0.135。
4.5.2 fMRI结果
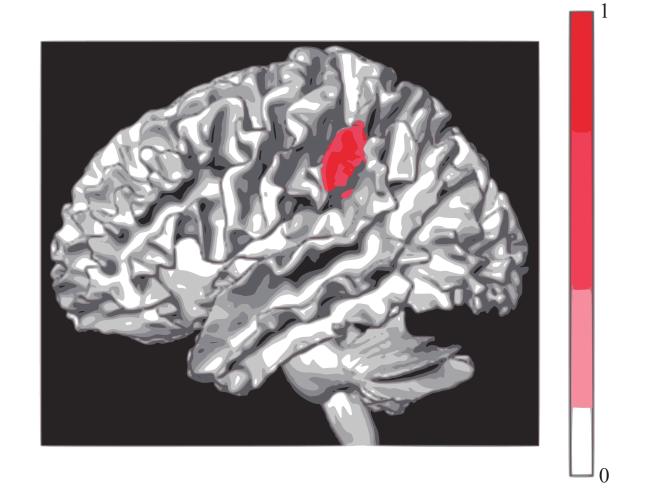
(1)全脑分析。结果显示不同组别在左侧IPL存在显著差异。具体而言,左侧IPL的激活体素数量为57,峰值点的MNI坐标为(−32, −48, 60),F值为12.10。该结果在体素水平未校正时显著(p<0.001),且在团块水平经过FDR校正后显著(p=0.023),即专家组在正确预判时的激活程度显著高于对照组(图3 )。
(2)基于ROI分析。在对速度结果进行预判的过程中,比较专家组和对照组兴趣区的信号值发现,在左侧IPL(−32, −48, 60)和右侧LOTC(36, −54, −14),专家组正确判断速度时的激活程度比对照组更强[左侧IPL: t(58)=2.61, p<0.001, Cohen’s d=0.96; 右侧LOTC: t(58)=2.33, p<0.001, Cohen’s d=0.86],而在其他兴趣区上均未发现显著的激活差异。在预判快速球时,专家组比对照组在左侧IPL和右侧LOTC均发现更强的激活[左侧IPL: t(58)=2.48, p<0.001, Cohen’s d=0.91; 右侧LOTC: t(58)=2.32, p<0.001, Cohen’s d=0.86],在预判慢速球时发现左侧IPL和右侧LOTC上专家组的激活更强[左侧IPL: t(58)=2.49, p<0.001, Cohen’s d=0.92; 右侧LOTC: t(58)=2.27, p<0.001, Cohen’s d=0.83]。在对专家组和对照组两个群体分别进行配对样本t检验,结果发现专家组在右侧的背外侧前额叶区域,预判快速的激活程度显著高于预判慢速球,t(58)=−2.60,p<0.001,Cohen’s d=0.79,其他区域没有发现显著的激活差异,而对照组在判断两种速度球时均未发现有任何兴趣区存在显著差异。
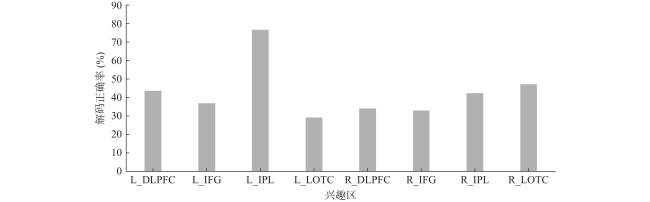
(3)MVPA结果。结果显示,专家组在左侧IPL的分类准确性达到76.7%,p=0.016(图4 ),其他特征区域则未达到随机水平(50%),而对照组所有特征区域均未达到随机水平(50%)。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
4.6 讨论
在fMRI结果上,专家组左侧IPL和右侧LOTC的激活更强,并且左侧IPL可能是表征快速和慢速球的关键脑区。这一结果符合了前期的假设,IPL是镜像神经系统的主要节点,也是理解动作意图的关键脑区(Wright et al., 2011, 2013),提示了速度预判在动作理解的层级加工中属于高层级的意图理解范畴。
5 总讨论
实验1首先验证了运动专家利用观察到的动作的视觉信息,能够更有效地进行速度预判。基于模拟理论的观点,有运动专家能够更快速且精准地提取运动学信息,借助内部模型进行动作模拟,进而能够更好地预判动作的结果(Colling et al., 2014; Kilner et al., 2007)。大量前人的研究从对空间的预判任务上验证了这一理论,比如乒乓球运动员能更准地预判发球的落点(Wang et al., 2019)。而当动作不具有时间连贯性时,专家组和对照组的正确率都会显著下降。这可能是因为在“不一致”条件下呈现的信息不符合个体既有的知觉预期。对于专家组而言,这种信息违背了其基于长期训练形成的感觉运动经验,可能激活更复杂的认知加工过程,如动作意图的理解、模拟偏差的识别与调整等,因此在面对冲突信息时产生更多认知负荷,影响其判断准确性。相较之下,对照组缺乏系统的运动经验,可能更依赖于即时的表面特征进行判断,当时间连贯性被打破时,也会因缺乏稳定的策略而出现表现下降。总之,实验1的结果支持了模拟理论,说明运动经验不仅能提高空间预判的准确性,还能在速度预判加工中展现专家优势。
实验2结果显示,与完整动作相比,单独呈现助跑或摆腿信息均显著降低了速度预判的正确率。这一发现与前人采用时间阻断技术的研究结果一致(黎晓丹 等, 2018),表明完整动作提供的运动学信息有助于提升预判表现。值得注意的是,虽然助跑和摆腿信息对预判正确率的影响程度相近,但基于摆腿信息的加工时间显著更长。研究发现,运动学信息在足球预判中起关键作用:点球方向预判主要依赖摆腿时的髋关节角度等运动学信息(Abdelkader et al., 2021),而传球时机预判则更多取决于对传球意图的理解(Pedullà et al., 2020)。研究表明,守门员通过整合助跑速度、支撑腿位置及射门者躯干方向等运动学信息,可显著提升点球防守成功率(Ridderinkhof et al., 2022)。值得注意的是,虽然摆腿和助跑信息对速度预判的贡献度相当,但其加工机制可能存在差异:摆腿信息涉及更深层的意图加工,而助跑信息则主要依赖速度感知。
实验3揭示了速度预判的神经机制:左侧IPL和右侧LOTC是速度预判的关键脑区,且专家组表现出更强的激活。具体而言,左侧IPL不仅参与区分快速和慢速踢球动作,还可能与时间预测网络协同工作,整合时间与距离信息进行速度预判(Coull & Nobre, 2008)。这一发现与Wright等人(2013)的研究一致,表明左侧IPL在熟练动作预判中起重要作用。除此之外,左侧IPL同样也会参与理解抓握动作和抓握目标时的加工过程(Wurm & Lingnau, 2015)。本研究推测左侧IPL可能通过编码运动学信息和理解动作意图来支持专家的速度预判。
另外,LOTC的显著激活可能说明本研究的速度预判任务可能涉及对速度概念性的理解,这或许是由于运动员在真实的训练和比赛情境中,对速度结果的预判通常代表了对战术策略的判断而非对单纯速度进行分类,即这一过程更多是理解对方的策略性任务来做出自身的反应,因此能够激活具有理解概念性知识的脑区。LOTC通常加工处理与身体感知相关的信息以及表征动作概念等,并且LOTC中所编码的运动信息是参与决策过程的一部分(Sitnikova et al., 2014)。
总而言之,对运动动作的速度预判需要加工运动学信息,包含对动作意图的高层级加工。在过去研究中发现,顶叶区对于识别不同形式的动作,联系视觉运动层次和动作目标层次起着重要作用(Wurm & Caramazza, 2019),本研究结果也发现左侧IPL能够分辨不同速度的点球,可能说明人们会将不同速度的点球定义为不同形式的动作加以识别与编码。
本研究的局限性主要包括缺少预判日常动作的对照,不能说明经验对于一般的动作是否有同样的影响,未来的研究可以探究不同人群在有共同经验的日常动作时,是否存在对速度预判加工的差异。另外,未来研究可以增加试次数量,以进一步提高MVPA的分类有效性。
6 结论
足球运动员对点球动作的速度预判具有专家优势,IPL和LOTC主要参与速度预判的加工。这种专家优势主要来源于对动作的速度预判的意图理解高层级加工。