
1 引言
网络游戏成瘾(internet gaming disorder, IGD)以过度和反复的游戏行为为核心特征,表现为对游戏行为的失控及戒断后的身心不适(American Psychiatric Association, 2013)。当下IGD的认知缺陷机制尚存在争议,尤其是在区分目标和干扰方面。值得注意的是,现有研究多依赖Stroop任务考察IGD的干扰控制,忽视了不同类型冲突的差异化机制。Stroop任务的冲突源于颜色和词义,难以模拟IGD真实情境中面临的冲突干扰。IGD的干扰控制是否会因冲突类型不同而不同?本研究通过整体−局部冲突任务,旨在回答:(1)游戏中长期的多目标处理经历是否会导致IGD在整体−局部冲突任务上表现出特异性加工?(2)这种加工的潜在神经机制是什么?
以往关于IGD干扰控制的研究主要基于Stroop范式。例如,Nasser等人(2021)招募IGD被试让其完成经典Stroop任务,同时记录被试完成任务时的大脑活动。结果发现,相较于对照组,IGD在完成任务时的正确率和反应时都更差,在前额叶有更小的脑激活,表明其在Stroop任务中存在缺陷。与此不同,Wu等人(2020)同样让IGD和对照组完成经典Stroop任务,结果却发现两组被试在任务表现上没有显著差异。近期研究者使用fMRI技术考察问题性游戏玩家在完成经典Stroop任务时的大脑活动特点,结果发现两组被试在任务正确率和反应时上没有显著差异,但是在大脑激活上,相较于对照组,问题性游戏玩家在背外侧前额叶有更大的激活(Shen et al., 2023)。在成瘾Stroop任务上,研究者发现相较于对照组,IGD组对电脑相关词的反应时更长,表现出对这些词的注意偏向(Jeromin et al., 2016a)。而在Jeromin等人(2016b)的另一项研究中,同样使用成瘾Stroop任务,结果却发现IGD组和对照组对电脑相关词和中性词的反应时间没有显著差异。
在经典Stroop任务中,冲突干扰源于词的语义和颜色,该任务主要考察认知控制能力,侧重自上而下加工(Ehlis et al., 2024; Gerlach & Poirel, 2020)。成瘾Stroop任务使用成瘾相关词作为材料,冲突干扰源于成瘾刺激和任务目标,该任务主要考察成瘾相关刺激引发的注意偏向和情绪干扰(杨玲 等, 2013; Jeromin et al., 2016a)。尽管上述两个任务能一定程度考察IGD的认知控制和注意偏向问题,但冲突干扰形式均难以模拟IGD在真实情境中面临的冲突干扰,可解释性和推广性低,这可能是研究结果不一致的原因。游戏场景中,玩家通常在多层级(全局和局部)目标之间动态切换,例如同时处理宏观战场信息(整体)与微观个人操作(局部)。而Stroop任务主要以词的颜色−语义冲突为主,这导致该任务难以捕捉IGD个体在层级冲突之间的认知加工特点(Toth et al., 2019; Wiley et al., 2024)。相反,整体−局部任务侧重被试在整体目标与局部目标之间的切换,能很好地弥补Stroop任务的局限,进而反映IGD在层级冲突之间的加工特点。
I-PACE模型指出,游戏成瘾是个体、环境和游戏机制交互影响的结果(Brand et al., 2025),目前很多学者也强调应该关注玩家在更贴合真实游戏环境下的行为特征,这些特征能更可靠地预测IGD(Zhang et al., 2025; Zhang et al., 2023)。依据上述观点,对于IGD来讲,在众多干扰刺激中锁定目标,以及在整体−局部刺激中灵活切换更切合这一群体的现实情境,在这些情境下的加工特点能够更深入地理解IGD。IGD在整体−局部冲突任务中的表现如何?任务表现背后的神经机制是什么?这些问题尚不清楚。因此,本研究拟让IGD完成整体−局部冲突任务,同时记录被试在这一过程中的大脑信号,以期回答上述两个问题。研究结果有望拓宽IGD干扰控制领域,加深对IGD在整体−局部冲突任务中行为特点和神经机制的认识。
传统行为指标在揭示认知加工上可能敏感性不够,且难以探讨内在神经机制。基于此,本研究引入功能性近红外光谱脑成像技术(functional near-infrared spectroscopy, fNIRS),fNIRS是一种非侵入性的神经成像技术,能让研究者在复杂的实验任务中实时测量被试的神经活动。此外该技术对于运动耐受性更强,所以被广泛应用于神经机制的研究(Li et al., 2023; Pinti et al., 2020)。现有文献多采用传统抑制控制范式(如go/no-go任务、停止信号任务和Stroop任务)考察IGD的抑制控制能力(Shen et al., 2023; Wang et al., 2021),而忽视了这一群体整体−局部的干扰控制能力。仅有的几项研究均是使用行为实验探讨游戏玩家在整体−局部任务上的加工特点,尚未有研究深入挖掘其背后的神经机制(Franceschini et al., 2022; Wong & Chang, 2018)。本研究首次结合整体−局部冲突任务与fNIRS技术,考察IGD个体在整体−局部任务中的神经机制。大脑前额叶(prefrontal cortex, PFC)通常被认为是执行功能的关键脑区,参与目标制定、注意加工、抑制干扰、决策等加工过程(Banich & Depue, 2015; Luijten et al., 2014; Munakata et al., 2011),干扰控制涉及对目标刺激的注意以及对干扰冲突的加工,这些与PFC紧密相关(Wu et al., 2020; Xu et al., 2024)。因此,本研究主要关注PFC区域,包括背外侧前额叶、眶额叶、额极等区域的激活变化。
综上,本研究采用整体−局部冲突任务结合fNIRS技术考察IGD的干扰控制能力及其神经机制。研究发现以动作视频游戏为主的网络游戏可能会增强个体将注意力集中在目标并且忽略干扰信息的能力(Bavelier & Green, 2019)。在游戏过程中个体需要对游戏的细节加以注意,搜寻敌人将在哪里出现并对游戏变化进行快速响应(郭文欣 等, 2023)。结合激励敏化理论(incentive-sensitization theory),真实游戏情境中,玩家在整体−局部目标间频繁切换会导致大脑纹状体−前额叶的环路敏化(Berridge & Robinson, 2016),而这种敏化可能会增强个体在整体−局部任务上的表现。因此本研究假设,由于在游戏中长期的多目标处理和转换训练,IGD在整体−局部冲突任务上可能表现出特异性优势。具体表现为,IGD组在整体−局部冲突任务上的正确率更高,被试在背外侧前额叶的激活(β)更大。此外,在额极和眶额叶,IGD组在一致和不一致条件下激活(β)可能没有显著差异。且这些脑区的激活水平与被试的任务表现相关。
2 研究方法
2.1 被试
本研究使用游戏成瘾量表和网络游戏成瘾结构化访谈(Structured Clinical Interview for Internet Gaming Disorder, SCI-IGD)筛选被试。网络游戏成瘾的测量使用杨文娇和周治金(2004)编制的网络成瘾类型量表中的网络游戏成瘾问卷,采用李克特五点计分,得分越高表明网络游戏成瘾越严重,高于25分的个体被认为网络游戏成瘾,该工具在本研究中的Cronbach’s α为0.87。SCI-IGD临床访谈由Koo等人(2017)根据DSM-5中网络游戏成瘾的诊断标准制定,用于筛选网络游戏成瘾个体。满足9条标准中5条及以上的被认为网络游戏成瘾,该工具的LRP(likelihood ratio positive)为10.93,LRN(likelihood ratio negative)为0.35,表明其能很好地筛选出成瘾个体。此外,确保被试最近半年未服用过利他林等精神类药物,没有严重的情绪和精神健康问题。同时满足以上三个标准的被试才能参加正式实验。使用G*Power软件计算所需样本量,本研究为2(组别:IGD组、对照组)×2(加工水平:整体、局部)×3(冲突:一致、不一致、中性)的混合实验设计,在预测中等效应(ƒ=0.25)且显著性水平α=0.05时,如果要达到80%的统计检验力水平,每组需要10人,共20人。经过筛选一共50人参加本实验,6人数据因为不符合要求被剔除(近红外数据超一半通道的信号变异系数大于15%),最终纳入数据分析的被试共44人,其中IGD组22人(男生14人,女生8人),对照组22人(男生16人,女生6人)。被试的年龄范围在18~24岁,平均年龄为20.71±1.61岁,所有被试均为右利手。本研究通过了华中师范大学心理学院伦理委员会审核。
2.2 实验材料和流程
实验材料和流程见图1 。一个整体形状(菱形、正方形或圆形)由八个局部形状(菱形、正方形或圆形)构成。局部形状与整体形状可构成一致、不一致、中性三种材料类型,当形状在整体和局部上一致时(如由局部菱形组成的整体菱形)构成一致性材料;当形状在两个水平上不同时(如由局部正方形组成的整体菱形)构成不一致性材料;当某一个水平呈现中性形状(即圆形)并且在整体或局部呈现目标形状(即菱形或正方形)时构成中性材料。
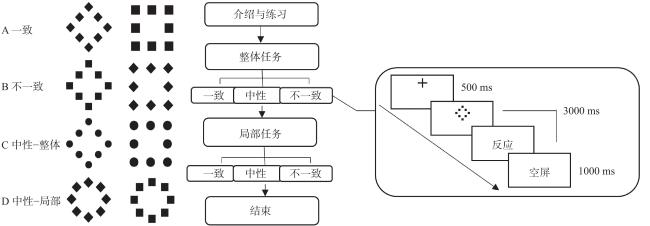
实验流程包括练习和正式实验,在进行正式实验前,被试需要进行练习,正确率达到80%以上才能参与正式实验。被试的任务是根据指导语对屏幕上呈现的刺激进行判断并做出相应的按键反应,当刺激整体或局部形状为菱形时按“Q”键,形状为正方形时按“P”键。在整体任务阶段,被试需要忽略刺激的局部形状根据整体形状进行判断;局部任务阶段,被试需要忽略刺激的整体形状根据局部形状进行判断。所有实验材料均在E-Prime软件上以白色背景呈现,并在屏幕的上、下、左、右4个位置随机出现。整体任务和局部任务各包含3个组块,分别对应一致、不一致和中性条件,每个组块40个试次,共240个试次。组块之间的呈现顺序随机,组块之间休息18 s以保证血氧恢复到基线水平。
2.3 近红外数据采集
本研究使用NIRSout系统实时记录被试在任务过程中氧合血红蛋白(HbO)、脱氧血红蛋白(HbR)和总血氧(HbT)的浓度变化。该系统使用785 nm和830 nm两种波长的近红外光,采样率为7.81 Hz。仪器探头包括8个光源和7个探测器,探头之间间隔3 cm,构成20个通道。光极排布参照国际10-20系统,覆盖前额叶,通道布局见图2 。使用NIRSout系统配置的3D定位笔进行定位,并通过NIRS_SPM软件采用概率配准法将各个位置与MNI空间坐标进行配准,确定每个通道与布鲁德曼分区(Brodmann areas, BA)之间的对应关系,结果见表1 。
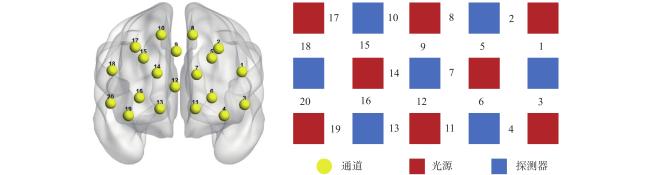
表1 通道对应脑区及覆盖率 |
| 通道 | MNI坐标 | BA分区 | 覆盖率(%) | ||
| X | Y | Z | |||
| CH1 | −46 | 45 | 26 | 45-布洛卡三角区 | 70 |
| CH2 | −28 | 45 | 43 | 9-背外侧前额叶 | 92 |
| CH3 | −47 | 52 | 3 | 46-背外侧前额叶 | 89 |
| CH4 | −34 | 65 | −6 | 10-额极区 | 38 |
| 11-眶额叶 | 42 | ||||
| CH5 | −41 | 56 | 17 | 46-背外侧前额叶 | 84 |
| CH6 | −24 | 70 | 8 | 10-额极区 | 83 |
| CH7 | −14 | 67 | 26 | 10-额极区 | 94 |
| CH8 | −10 | 47 | 52 | 9-背外侧前额叶 | 78 |
| CH9 | 2 | 55 | 42 | 9-背外侧前额叶 | 97 |
| CH10 | 13 | 46 | 53 | 9-背外侧前额叶 | 74 |
| CH11 | −11 | 73 | −2 | 10-额极区 | 62 |
| 11-眶额叶 | 38 | ||||
| CH12 | 3 | 69 | 17 | 10-额极区 | 100 |
| CH13 | 16 | 73 | −1 | 10-额极区 | 57 |
| 11-眶额叶 | 43 | ||||
| CH14 | 17 | 67 | 26 | 10-额极区 | 95 |
| CH15 | 29 | 69 | 8 | 10-额极区 | 82 |
| CH16 | 45 | 54 | 19 | 46-背外侧前额叶 | 84 |
| CH17 | 33 | 44 | 44 | 9-背外侧前额叶 | 94 |
| CH18 | 49 | 42 | 27 | 45-布洛卡三角区 | 88 |
| CH19 | 38 | 65 | −6 | 10-额极区 | 41 |
| 11-眶额叶 | 35 | ||||
| CH20 | 51 | 49 | 4 | 46-背外侧前额叶 | 79 |
注:仅报告覆盖率≥30%的脑区。 |
2.4 数据分析
使用R语言(版本4.3.3)对被试在任务中的正确率和反应时进行2(组别:IGD组、对照组)×2(加工水平:整体、局部)×3(冲突:一致、不一致、中性)的混合方差分析以及行为数据和近红外数据的相关分析。使用nirsLAB软件对近红外数据进行分析,具体包括:(1)裁切掉实验前后与任务无关的数据;(2)使用样条插值法去除尖峰伪迹;(3)使用0.01~0.2 Hz的带通滤波去除运动和生理伪迹;(4)基于一般线性模型计算每个被试在不同条件下的β值,将其作为脑激活的指标。研究发现氧合血红蛋白(HbO)对任务刺激的变化更加敏感(杨海波 等, 2019; Hoshi et al., 2001),因此后续分析以HbO变化的β值作为因变量指标进行混合方差分析。实验结果在通道间采用FDR方法对p值进行校正,最后使用BrainNet Viewer软件对脑成像结果进行可视化。
3 结果
3.1 行为结果
两组被试在任务中的正确率和反应时见表2 。混合方差分析发现,在正确率上,冲突主效应显著,F(2, 84)=5.54,p=0.014,η$ {}_{\mathrm{p}}^{2} $ =0.12,不一致条件下的正确率(M=0.96, SD=0.05)显著低于一致条件下的正确率(M=0.98, SD=0.03)。组别和冲突的交互效应显著,F(2, 84)=3.87,p=0.042,η$ {}_{\mathrm{p}}^{2} $ =0.08,简单效应分析发现,对照组在不一致条件下的正确率(M=0.94, SD=0.11)显著低于一致条件下(M=0.99, SD=0.02),而IGD组在两种条件下的正确率没有显著差异。在反应时上,加工水平主效应显著,F(1, 42)=6.60,p=0.014,η$ {}_{\mathrm{p}}^{2} $ =0.14,局部条件下的反应时(M=551 ms, SD=104 ms)显著长于整体条件下的反应时(M=531 ms, SD=96 ms)。冲突主效应显著,F(2, 84)=31.17,p<0.001,η$ {}_{\mathrm{p}}^{2} $ =0.43,不一致条件下的反应时(M=573 ms, SD=91 ms)显著长于一致(M=514 ms, SD=77 ms)和中性条件下的反应时(M=535 ms, SD=84 ms),中性条件下的反应时(M=535 ms, SD=84 ms)显著长于一致条件下的反应时(M=514 ms, SD=77 ms)。
表2 两组被试在不同条件下的正确率和反应时(M±SD) |
| 整体 | 局部 | |||||||
| 一致 | 不一致 | 中性 | 一致 | 不一致 | 中性 | |||
| ACC | IGD组 | 0.96±0.04 | 0.97±0.03 | 0.97±0.04 | 0.99±0.02 | 0.98±0.02 | 0.98±0.02 | |
| 对照组 | 0.99±0.02 | 0.94±0.11 | 0.98±0.03 | 0.98±0.02 | 0.96±0.04 | 0.98±0.03 | ||
| RT(ms) | IGD组 | 492±68 | 532±67 | 511±74 | 507±71 | 571±68 | 540±71 | |
| 对照组 | 520±106 | 600±148 | 528±116 | 538±63 | 591±82 | 558±77 | ||
3.2 近红外结果
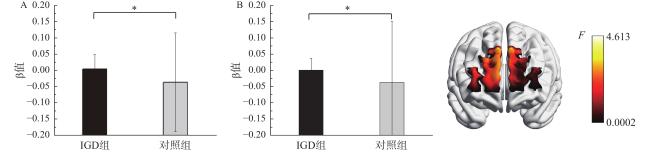
以大脑激活指标β值为因变量,进行2(组别:IGD组、对照组)×2(加工水平:整体、局部)×3(冲突:一致、不一致、中性)混合方差分析。结果发现,通道8和通道10组别主效应显著,F(1, 42)=4.41,p=0.042,η$ {}_{\mathrm{p}}^{2} $ =0.10,F(1, 42)=4.61,p=0.038,η$ {}_{\mathrm{p}}^{2} $ =0.10,IGD组的激活水平显著强于对照组,见图3 。通道8和通道10均对应背外侧前额叶。
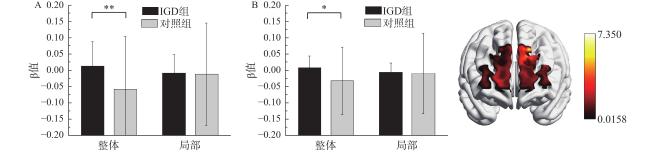
在通道2和通道13上,组别与加工水平的交互效应显著,F(1, 42)=7.35,p=0.010,η$ {}_{\mathrm{p}}^{2} $ =0.15,F(1, 42)=5.04,p=0.030,η$ {}_{\mathrm{p}}^{2} $ =0.11。简单效应分析发现,当加工水平为整体时,IGD组的激活水平显著高于对照组,当加工水平为局部时,两组的激活水平没有显著差异,见图4 。通道2对应背外侧前额叶,通道13对应额极。
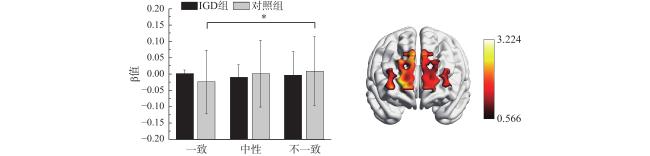
在通道19上,组别与冲突的交互效应显著,F(2, 84)=3.22,p=0.045,η$ {}_{\mathrm{p}}^{2} $ =0.07。简单效应分析发现,对照组在不一致条件下的激活显著高于一致条件下,而IGD组在三个条件下的激活水平没有显著差异,见图5 。该通道对应额极和眶额叶。其余效应均不显著(ps>0.05)。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3.3 相关分析结果
对各个条件下差异显著通道的β值与行为结果之间进行相关分析。结果发现,在局部一致条件下,被试的任务正确率与通道2、8、10的激活强度显著正相关(ps<0.05)。在局部不一致条件下,被试的任务正确率与通道19的激活强度显著负相关(p<0.001)。上述结果表明,在局部一致条件下(没有冲突时),背外侧前额叶激活越高,被试任务正确率越高,而在局部不一致条件下(有冲突时),额极和眶额叶激活越低,被试任务正确率越高,见表3 。
表3 差异显著通道激活强度与行为结果的相关分析 |
| 2 | 8 | 10 | 13 | 19 | ||||||||||
| r | p | r | p | r | p | r | p | r | p | |||||
| 整体一致-ACC | 0.00 | 0.978 | 0.03 | 0.829 | 0.01 | 0.971 | −0.11 | 0.475 | −0.12 | 0.426 | ||||
| 整体不一致-ACC | −0.03 | 0.860 | −0.01 | 0.949 | −0.00 | 0.949 | 0.05 | 0.765 | 0.03 | 0.834 | ||||
| 整体中性-ACC | 0.26 | 0.083 | 0.23 | 0.132 | 0.23 | 0.132 | −0.04 | 0.804 | −0.16 | 0.308 | ||||
| 局部一致-ACC | 0.42** | 0.005 | 0.36* | 0.015 | 0.39** | 0.008 | 0.12 | 0.450 | 0.07 | 0.675 | ||||
| 局部不一致-ACC | −0.04 | 0.778 | −0.03 | 0.870 | −0.00 | 0.995 | −0.17 | 0.277 | −0.51*** | <0.001 | ||||
| 局部中性-ACC | −0.12 | 0.443 | −0.18 | 0.254 | −0.20 | 0.185 | 0.00 | 0.993 | −0.14 | 0.382 | ||||
| 整体一致-RT | 0.15 | 0.340 | 0.17 | 0.280 | 0.12 | 0.455 | 0.12 | 0.439 | 0.06 | 0.676 | ||||
| 整体不一致-RT | 0.05 | 0.730 | 0.02 | 0.879 | −0.03 | 0.831 | 0.09 | 0.544 | 0.06 | 0.706 | ||||
| 整体中性-RT | −0.06 | 0.696 | 0.06 | 0.700 | 0.05 | 0.740 | 0.14 | 0.352 | 0.04 | 0.819 | ||||
| 局部一致-RT | −0.04 | 0.813 | 0.12 | 0.452 | 0.12 | 0.452 | 0.12 | 0.438 | 0.11 | 0.493 | ||||
| 局部不一致-RT | −0.01 | 0.969 | 0.13 | 0.415 | 0.09 | 0.552 | −0.08 | 0.584 | 0.06 | 0.702 | ||||
| 局部中性-RT | −0.03 | 0.848 | 0.12 | 0.419 | −0.04 | 0.817 | −0.04 | 0.779 | 0.01 | 0.935 | ||||
4 讨论
先前研究主要采用Stroop任务考察IGD自上而下的认知控制能力,然而针对该群体在自下而上干扰控制任务中的研究较为匮乏。为此,本研究采用整体−局部冲突任务结合fNIRS技术,系统考察了IGD的干扰控制特点。研究结果表明,IGD在整体−局部冲突任务中表现出加工优势。具体而言,局部不涉及冲突时,背外侧前额叶的激活介导了IGD更高的认知投入,从而提高了任务表现;局部涉及冲突时,额极和眶额叶的激活强度保持稳定,可能说明IGD对冲突的适应和自动化加工,进而促进了任务表现。本研究不仅拓展了IGD在干扰控制任务中的认知加工特点,还为其神经机制提供了新的实验证据,加深了对IGD群体的全面认识。
首先,行为结果发现,被试在不一致条件下的正确率显著低于一致条件,反应时则显著长于一致条件,此外,被试在局部条件下的反应时显著长于整体条件下的反应时,这些结果表明实验对加工水平和冲突的操纵有效,与以往研究一致(Gerlach & Poirel, 2020; Hayward et al., 2018)。值得注意的是,本研究发现对照组在不一致条件下的正确率显著低于一致条件,而IGD组在两种条件下的正确率没有显著差异。这一结果表明IGD群体并没有因为冲突的存在导致任务表现变差,相反IGD在应对整体−局部冲突时表现更好。不一致条件通常需要被试投入更多的认知资源,对于未受过训练的人在该条件下的表现普遍更差,表现出整体加工优势(Gerlach & Poirel, 2020; Weinbach & Henik, 2014)。而IGD组在一致和不一致条件下的正确率没有显著差异,说明其在整体−局部冲突任务中存在加工优势。双过程理论指出人的认知加工存在两套系统,系统一是自动化的、不受认知资源限制,多用于加工自下而上的信息,系统二是意识控制的、受认知资源的限制,多用于加工自上而下的信息(Evans, 2017; Fleming & Bartholow, 2014)。IGD由于长期玩电子游戏,游戏场景的复杂性和目标的频繁切换,要求玩家对不同位置、不同类型的刺激快速响应(Griffiths & Nuyens, 2017; King et al., 2010),这种长期的训练可能使其对冲突的加工更加敏感并能快速适应冲突,甚至发展为自动化加工,这可能是IGD在整体−局部冲突任务上的表现更好的原因之一。近几年也有研究者发现,玩电子游戏可以提高个体的注意广度(Argilés et al., 2023)和注意控制(Bertoni et al., 2021)。
不涉及冲突时,背外侧前额叶与额极的激活可能反映了被试的干扰控制能力。近红外结果发现,在背外侧前额叶,IGD组的激活显著高于对照组。此外,当加工水平为整体时,IGD组在背外侧前额叶的激活显著高于对照组,当加工水平为局部时,两组激活没有显著差异,在额极区域也发现相似结果。此外,相关分析发现,局部一致条件下背外侧前额叶的激活强度与任务正确率显著正相关。背外侧前额叶是参与执行控制的关键脑区,负责冲突监测和冲突解决(Friedman & Robbins, 2022; Lee & Kim, 2019)。近期Shen等人(2023)探讨了问题性游戏玩家在完成Stroop任务时的大脑神经活动特点,结果发现,相较于对照组,问题性游戏玩家在完成任务时背外侧前额叶的激活更强,本研究结果与其一致。上述结果表明,当不涉及冲突时,背外侧前额叶的激活可能介导了IGD群体更高的认知投入,从而提高了任务表现。
冲突条件下,额极和眶额叶的激活模式可能反映了被试对冲突的适应和自动化加工程度。本研究发现,在额极和眶额叶,对照组在不一致条件下的激活大于一致条件,而IGD组在两个条件下的激活没有显著差异,与本研究的行为结果一致。相关分析发现,在局部不一致条件下(即冲突条件),额极和眶额叶的激活强度与任务正确率显著负相关。研究发现人类和猕猴的额极在对抑制控制任务的适应与调整中起着关键作用(Feizpour et al., 2024),华艳等人(2020)发现眶额叶皮层是自动情绪注意选择相关的重要脑区。上述发现表明,额极和眶额叶可能和对任务的适应和自动化加工有关。在本研究中,IGD面对冲突时,无需显著提高额极和眶额叶的激活水平就能很好地完成任务,这一现象可通过激励敏化理论解释(Berridge & Robinson, 2016)。长期在复杂的游戏场景中快速锁定目标并灵活切换可能引发了中脑边缘系统的神经适应性敏化,促使IGD对整体−局部冲突更敏感。敏化的纹状体−额叶环路可能通过强化自下而上的加工,驱动个体无需大幅提高额叶激活水平就能高效解决冲突,这与双过程理论中系统一的自动化加工一致(Evans, 2017; Fleming & Bartholow, 2014)。因此,IGD能在额极和眶额叶激活没有显著提高的情况下很好地应对冲突任务,表现出整体−局部加工优势。
本研究通过整体−局部冲突任务并结合fNIRS技术,系统考察了IGD的干扰控制特点及其神经机制。研究发现IGD在整体−局部冲突任务中表现出加工优势。不涉及冲突时,背外侧前额叶的激活介导了IGD更高的认知投入,从而提高了任务表现;涉及冲突时,额极和眶额叶的激活强度保持稳定,可能说明IGD对冲突的适应和自动化加工,进而促进了任务表现。与以往研究相比,本研究首次揭示了IGD在自下而上整体−局部冲突任务中的独特优势,并发现背外侧前额叶、额极和眶额叶在这一过程中可能发挥着关键作用。这一发现不仅加深了人们对IGD群体的全面认识,还为未来研究提供了新方向。例如,未来可进一步探讨IGD在其他干扰任务中的表现,验证这一加工优势是否具有跨任务的普遍性。本研究的局限需要被关注,由于fNIRS自身局限,被试在完成任务时大脑深层区域的变化无法捕获,未来可结合fMRI进一步考察大脑深层区域的变化特点。
5 结论
IGD在整体−局部冲突任务中表现出加工优势。在不涉及冲突时,背外侧前额叶的激活可能介导了IGD更高的认知投入,从而提高了任务表现;在涉及冲突时,额极和眶额叶的激活强度保持稳定,可能说明IGD对冲突的适应和自动化加工,进而促进任务表现。